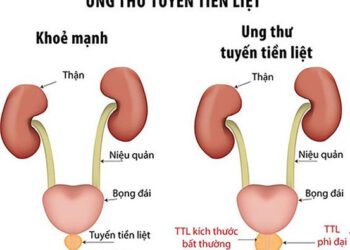
Điều trị ung thư tuyến tiền liệt bằng thuốc nam
Mời xem toàn văn báo cáo tại đây: Thuoc-nam-trong-ho-tro-ung-thu-tuyen-tien-liet
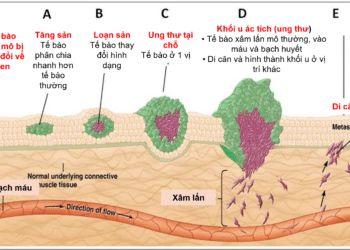
Thuốc đông – nam dược với bệnh ung thư
Mời xem toàn văn báo cáo tại đây: Hỗ trợ điều trị ung thư bằng thuốc Đông - Nam dược
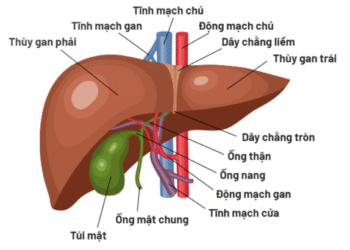
Hỗ trợ điều trị ung thư gan bằng thuốc Nam
Mời xem toàn văn báo cáo tại đường link sau: Thuốc Nam trong hỗ trợ điều trị ung thư gan




























NGHIÊN CỨU
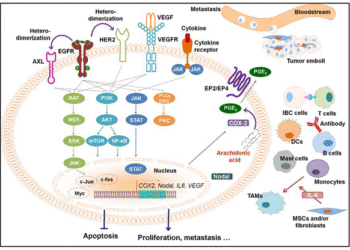


ƯNG THƯ

Một số loại rau củ có tác dụng hỗ trợ chữa ung thư.
Lời bạt: Nhóm biên soạn đưa những thông tin dưới đây, người mắc loại ung thư khác nhau, nguy cơ...
Read more
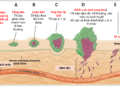
TIM MẠCH

BÀI MỚI

Một số loại rau củ có tác dụng hỗ trợ chữa ung thư.
Lời bạt: Nhóm biên soạn đưa những thông tin dưới đây, người mắc loại ung thư khác nhau, nguy cơ...
Điều trị ung thư tuyến tiền liệt bằng thuốc nam
Mời xem toàn văn báo cáo tại đây: Thuoc-nam-trong-ho-tro-ung-thu-tuyen-tien-liet
Thuốc đông – nam dược với bệnh ung thư
Mời xem toàn văn báo cáo tại đây: Hỗ trợ điều trị ung thư bằng thuốc Đông - Nam dược
Hỗ trợ điều trị ung thư gan bằng thuốc Nam
Mời xem toàn văn báo cáo tại đường link sau: Thuốc Nam trong hỗ trợ điều trị ung thư gan
Thuốc Nam trong hỗ trợ điều trị ung thư phổi
Mời xem toàn văn báo cáo tại đường link sau: Thuốc Nam hỗ trợ điều trị ung thư phổi
Thuốc Nam trong hỗ trợ điều trị ung thư thực quản
Mời xem toàn văn báo cáo tại đường link: Thuốc Nam hỗ trợ điều trị ung thư thực quản

Nghệ đen – Dược liệu tiềm năng chống ung thư phổi và tuyến tiền liệt
Nghệ đen (Curcuma zedoaria) hay còn gọi là Nga truật là một loại dược liệu thuộc họ Gừng (Zingiberaceae), được...

Cúc thơm – Tiềm năng điều trị u đa tủy xương
Bệnh đa u tủy xương là một dạng ung thư máu nguy hiểm, khi các tế bào tương bào ác...

Tảo nâu – Tiềm năng điều trị u limpho
Tảo nâu Nhật Bản (Cladosiphon okamuranus Tokida), còn gọi là Mozuku, là một loại tảo biển thuộc họ Chordariaceae, phân...

Hạt bông
U đa tủy xương là một loại ung thư máu ác tính, phát sinh từ tế bào plasma trong tủy...
XEM NHIỀU NHẤT
-

Tìm hiểu trường sinh học để hiểu thêm về tâm linh, ngoại cảm, thần giao cách cảm, chữa bệnh từ xa… và một số điều trị “thần bí” khác (2)
-
Tự chăm sóc và khuyến cáo người cao tuổi trong mùa đông giá lạnh
-
Vượt qua cái chết chỉ với ba cây thuốc nam
-
Khung chậu sản khoa
-
Một số loại rau củ có tác dụng hỗ trợ chữa ung thư.
-
Thảo dược với hẹp mạch vành theo dòng thời gian
-
Chè Shan Tuyết – đặc biệt với khả năng chống ung thư
-
Tuyến Cận Giáp
-
Kích thích âm vật: truy tìm liên hệ thần kinh âm vật – cột sống
-
Chữa loãng xương không cần cung cấp bổ sung canxi

{kind=link}
{kind=link}