
Chè shan tuyết – Tả Phìn Hồ
Chè Shan Tuyết được trồng trên vùng núi Tả Phìn Hồ thuộc Viện Y học bản địa Việt Nam, búp chè...

BÁC SĨ HOÀNG SẦM – NGƯỜI KHAI SƠN PHÁ THẠCH VÌ MỘT VÙNG ĐẤT
Có những người chọn ánh đèn đô thị. Có những người chọn ánh sáng từ bên trong – lặng lẽ,...
Những hiểu biết cơ bản về thiếu máu bất sản tủy
Lời bạt: Tôi có Anh bạn vong niên sinh 1948 chẳng may mắc bệnh thiểu máu bất sản tuỷ, thường...




























NGHIÊN CỨU

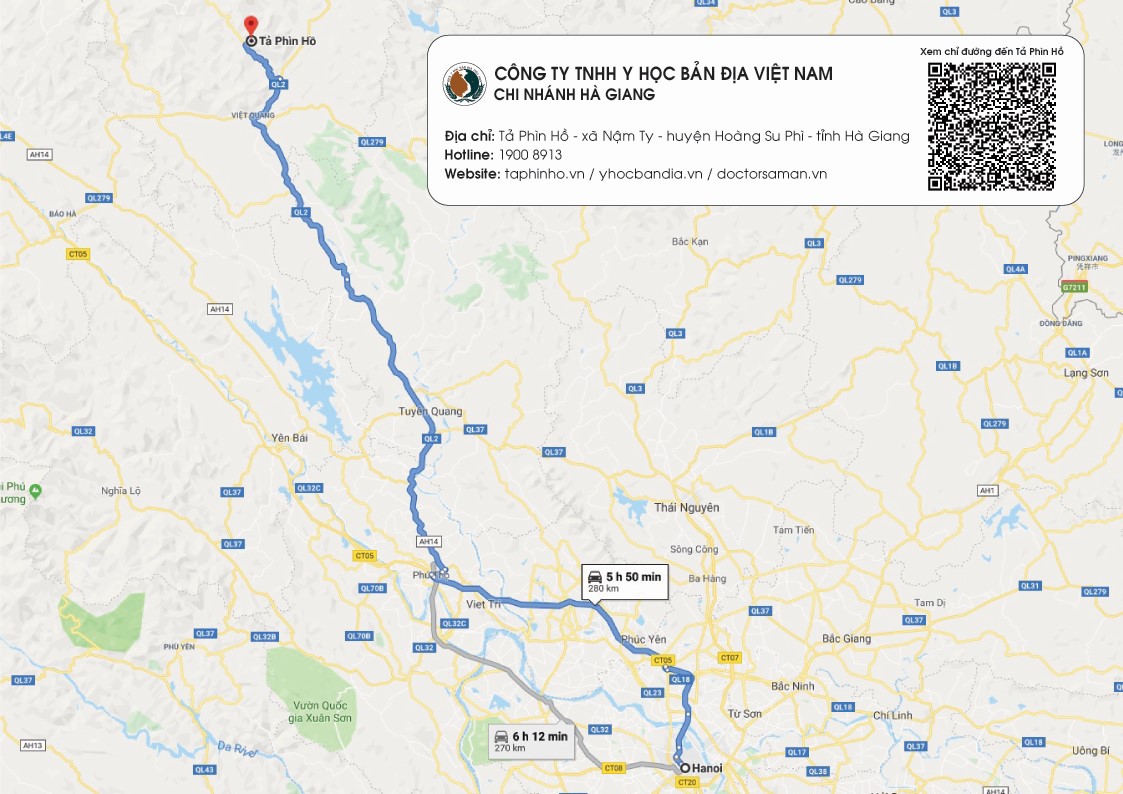
ƯNG THƯ
Tadalafil – Cơ hội mới trong điều hòa miễn dịch ở ung thư biểu mô vảy cổ tử cung?
Biên soạn: Hoàng Sầm – Viện Y học bản địa Việt Nam. Ung thư cổ tử cung, đặc biệt là...
Read moreDetails

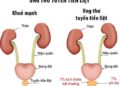
TIM MẠCH
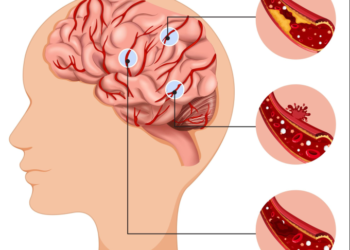

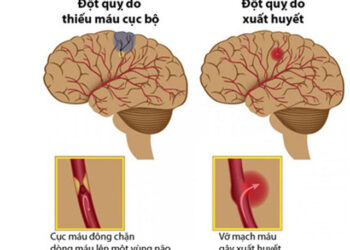
BÀI MỚI
GIỮ GÌN “ĐỘI SỬA GEN” – CÁCH PHÒNG UNG THƯ TỪ GỐC RỄ
Ở khắp vùng rẻo cao Tây Bắc – từ Tuần Giáo (Điện Biên), Thanh Sơn (Phú Thọ), đến tận Hà...
Chè shan tuyết – Tả Phìn Hồ
Chè Shan Tuyết được trồng trên vùng núi Tả Phìn Hồ thuộc Viện Y học bản địa Việt Nam, búp chè...
BÁC SĨ HOÀNG SẦM – NGƯỜI KHAI SƠN PHÁ THẠCH VÌ MỘT VÙNG ĐẤT
Có những người chọn ánh đèn đô thị. Có những người chọn ánh sáng từ bên trong – lặng lẽ,...
Những hiểu biết cơ bản về thiếu máu bất sản tủy
Lời bạt: Tôi có Anh bạn vong niên sinh 1948 chẳng may mắc bệnh thiểu máu bất sản tuỷ, thường...
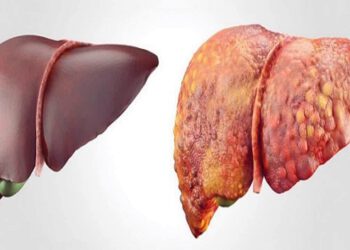
NHỮNG HIỂU BIẾT CƠ BẢN VỀ XƠ GAN VÀ CÁC ĐỊNH HƯỚNG ĐIỀU TRỊ
Lời bạt: cách đây 50 năm các tác giả hiểu về xơ gan đã đúng nhưng còn rất sơ sài,...
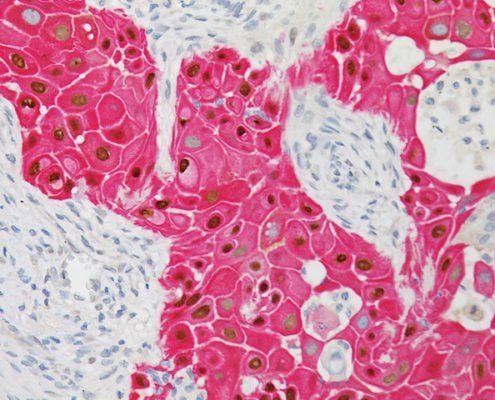
Tadalafil – Cơ hội mới trong điều hòa miễn dịch ở ung thư biểu mô vảy cổ tử cung?
Biên soạn: Hoàng Sầm – Viện Y học bản địa Việt Nam. Ung thư cổ tử cung, đặc biệt là...
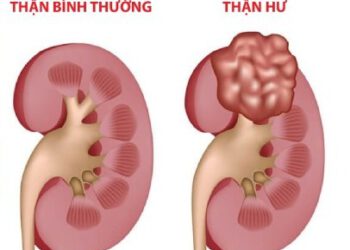
Sinh lý bệnh của Hội chứng thận hư
Lời bạt: Hiện nay Viện Y học bản địa Việt Nam đã và đang nghiên cứu lại cách chữa hội...

Tổng quan sinh lý học tóc và sắc tố melanin
I. CẤU TRÚC SINH HỌC CỦA NANG TÓC VÀ VAI TRÒ CỦA MELANOCYTE TRONG QUÁ TRÌNH SẮC TỐ HÓA Nang...
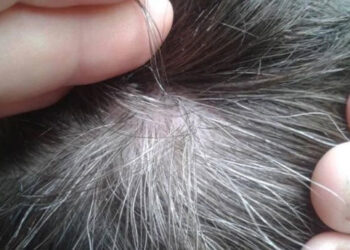
Bước đầu nghiên cứu bạc tóc và bạc tóc sớm
1. Giảm stress oxy hóa – Bảo vệ melanocyte 1.1 Tổng quan: Vai trò của stress oxy hóa trong bạc...

Thoái hóa chất trắng: Cơ chế bệnh sinh và cập nhật chiến lược điều trị dựa trên chứng cứ lâm sàng
Tóm tắt Thoái hóa chất trắng là quá trình bệnh lý phức tạp, liên quan đến nhiều cơ chế bệnh...
XEM NHIỀU NHẤT
-

Tìm hiểu trường sinh học để hiểu thêm về tâm linh, ngoại cảm, thần giao cách cảm, chữa bệnh từ xa… và một số điều trị “thần bí” khác (2)
-
Tự chăm sóc và khuyến cáo người cao tuổi trong mùa đông giá lạnh
-
Một số loại rau củ có tác dụng hỗ trợ chữa ung thư.
-
Khung chậu sản khoa
-
Vượt qua cái chết chỉ với ba cây thuốc nam
-
Những loại hoa quả có lợi cho người đang điều trị ung thư
-
GIỮ GÌN “ĐỘI SỬA GEN” – CÁCH PHÒNG UNG THƯ TỪ GỐC RỄ
-
Thảo dược với hẹp mạch vành theo dòng thời gian
-
Chè Shan Tuyết – đặc biệt với khả năng chống ung thư
-
Tuyến Cận Giáp

{kind=link}
{kind=link}